研究ハイライト
細胞死をもたらす新しい細胞融合現象の発見 〜受精後の花粉管誘引停止の全貌が明らかに〜
概要:
名古屋大学トランスフォーマティブ生命分子研究所(WPI-ITbM)の丸山 大輔 YLC特任助教および名古屋大学大学院理学研究科、JST戦略的創造研究推進事業ERATO東山ライブホロニクスプロジェクトの東山哲也教授(生殖分子情報学研究室)らは、 これまで例のない細胞同士の融合を伴う新しいタイプの細胞死を植物で発見しました。 雄しべの花粉が雌しべに付くと、花粉から伸び出した花粉管が、種子のもとになる組織に導かれます。 このとき花粉管を誘引する特殊な細胞が、卵細胞のとなりに2つある「助細胞」という細胞です。 しかし、受精後、役目を終えた助細胞の働きをいかに抑え、余分な花粉管の誘引を防ぐのかは、これまで明らかではありませんでした。 本研究グループは、受精後に残った方の助細胞の変化を、シロイヌナズナという植物で詳細に調べ、その結果、この助細胞が胚乳という細胞と融合して機能を失うことがわかりました。 植物の細胞は細胞壁で覆われていて普通は融合しないと考えられてきたことから、今回の現象は生物学の教科書を大きく書き変える新たな細胞融合の発見といえます。
研究について:
植物が種子をつくる際には、花粉から伸び出した花粉管が、種子のもとになる組織に導かれます。このとき花粉管を誘引する特殊な細胞が、卵細胞の隣に2つある「助細胞」という細胞です。しかし、余分な花粉管の誘引を防ぐため、役目を終えた助細胞の働きをいかに抑えるかは、これまで明らかではありませんでした。
花粉管が助細胞に到達すると、先端から2つの精細胞が勢いよく放出されます。これに伴い、助細胞が1つ潰れるとともに、精細胞の1つが卵細胞と受精して胚をつくりはじめ、もう1つの精細胞は中央細胞という大きな細胞と受精して胚乳という栄養組織を作り始めます。このとき残った方の助細胞に起きる変化を、名古屋大学の丸山大輔YLC特任助教と東山哲也教授らのグループは、シロイヌナズナという植物の助細胞に含まれるミトコンドリアなどを光らせて、レーザー顕微鏡で詳細に調べた結果、助細胞と胚乳が融合して互いの中身が混じり合う様子が観察されました。細胞が融合することは、電子顕微鏡による観察でも確認されました。この助細胞と胚乳の融合により、助細胞が含む誘引物質は急速に薄まります。さらに、助細胞の核が変性し、胚乳の核が分裂増殖していく過程で崩壊していく様子も明らかとなりました。
本研究によって、花粉管の誘引停止が起きる驚くべき仕組みが明らかとなりました。植物の細胞は細胞壁をもつため、これまで普通は融合しないと考えられており、110年以上も前に示された卵細胞の受精と中央細胞の受精しか融合は知られていませんでした。今回の現象は、受精に次ぐ新たな細胞融合の発見です。この融合相手の細胞に細胞死をもたらす全く新しいタイプの細胞融合現象は、植物細胞に対する見方を大きく変えるだけでなく、細胞の新たな機能を提示しました。これらの点で本研究成果は教科書を書き換える発見といえます。植物の細胞でも、受精以外の細胞融合を引き起こすプログラムの存在が示されたことから、植物における新たな細胞融合技術の開発にもつながることが期待されます。
本研究成果は名古屋大学 トランスフォーマティブ生命分子研究所(WPI-ITbM)、科学技術振興機構(JST)戦略的創造研究推進事業ERATO「東山ライブホロニクスプロジェクト」の研究の一環として得られたもので、学術誌「Cell」のオンライン速報版で公開されました。
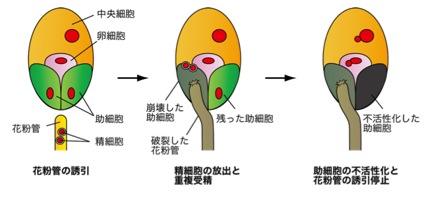
花粉が雌しべの柱頭に受粉すると、花粉は花粉管注1)を伸ばします。花粉管は雌しべ内部へと伸長して、雌しべの奥深くに存在する種子の元になる組織(胚珠注2))へと精細胞を届けます。このとき花粉管を正確に胚珠に導くために誘引物質を分泌するのが卵細胞の隣に2つある助細胞注3)です。花粉管が胚珠にたどり着くと(図1、左)、片方の助細胞の破壊とひきかえに内部にあった2つの精細胞を放出します(図1、中)。すると、精細胞の1つは卵細胞と受精して幼植物になる「胚」をつくり、他方は中央細胞と受精して胚への栄養を供給する細胞である「胚乳」をつくります。この重複受精注4)として知られる2つの受精の後で、生き残った方の助細胞は素早く不活性化されます(図1、右)。これによって、花粉管の誘引停止が起こるのですが、詳しいことは明らかではありませんでした。
名古屋大学の丸山大輔YLC特任助教と東山哲也教授らのグループは、受精後に残った方の助細胞の不活性化の謎に迫るため、シロイヌナズナという植物でミトコンドリアを緑色蛍光タンパク質のGFPでラベルした助細胞の経時観察を行いました。すると驚いたことに、助細胞のミトコンドリアが隣に位置する巨大な細胞である胚乳へと移動することがわかりました(図2・左)。さらに電子顕微鏡で助細胞と胚乳を隔てる部分を詳しく調べると、受精前にはみられなかった大きな穴が空いていることがわかりました(図2・右の矢印)。
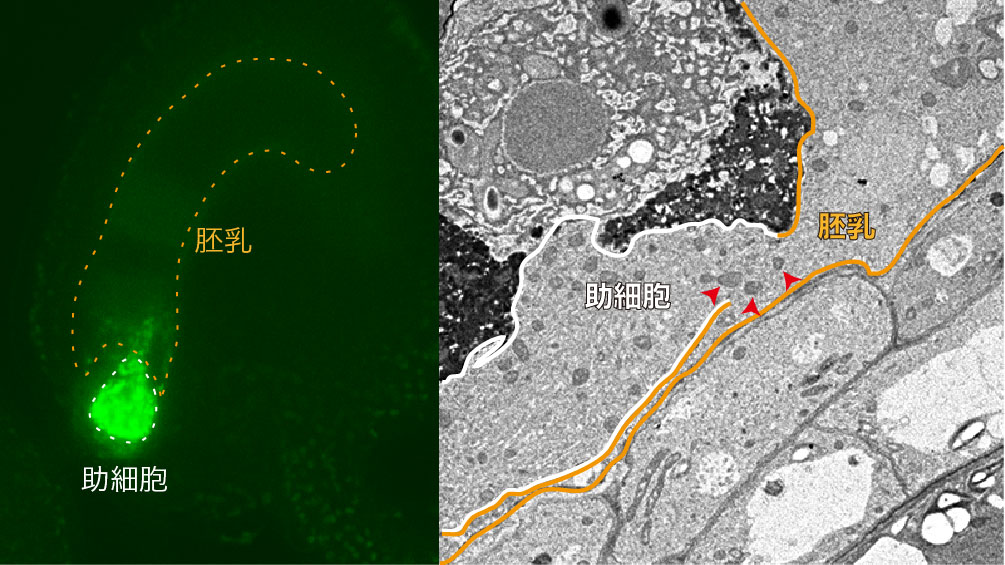
左図:助細胞のミトコンドリアを緑色蛍光タンパク質(GFP)で光らせた受精後の胚珠。白の破線で囲った助細胞から、オレンジの破線で囲った胚乳へと粒状構造をもつミトコンドリアが移動する様子をとらえることができました。 右図:受精後の胚珠の電子顕微鏡像。助細胞(白い実線)と胚乳(オレンジの実線)の間を隔てる細胞壁が一部で壊れており、その付近のミトコンドリア(矢尻)の移動を妨げる構造がないことがみてとれます。
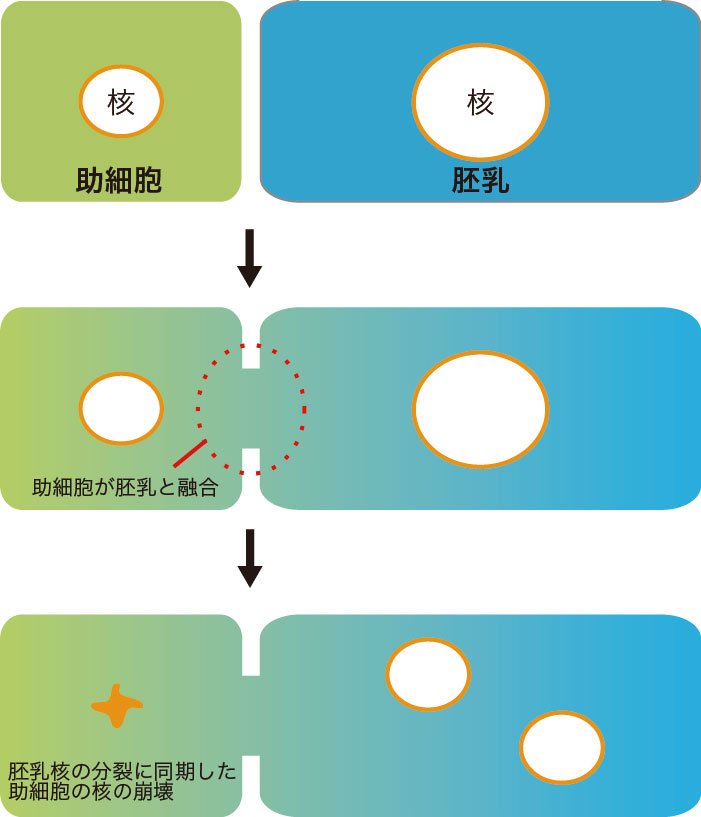
このため、受精後に助細胞と胚乳が融合し、助細胞の内容物が胚乳へと流出することがわかりました。移動するのはミトコンドリアだけではありません。助細胞では花粉管の誘引物質が絶えず作られて分泌されるため、花粉管が助細胞へと正確に伸長できると考えられています。この誘引物質をGFPでラベルした植物を観察すると、助細胞から胚乳へと分泌前の誘引物質が移動し、助細胞内での濃度が急激に低下しました。したがって、安定して分泌されていた誘引物質の供給が細胞融合によって途絶えることで2本目以降の花粉管は目標を失い、素早い花粉管の誘引停止が起きると考えられました。興味深いことに細胞融合は助細胞の核の運命にも関わっていました。融合後、助細胞の核は胚乳の核と共存する状態になっています。これらの核を経時観察すると、胚乳核の分裂に合わせ助細胞の核も分裂の準備に入るものの、最終的に分裂できず変性することがわかりました。つまり、細胞融合は誘引物質を含む細胞内容物の希釈と核の崩壊という二段階の仕組みで、助細胞を不活性化することがわかりました(図3)。
助細胞の速やかな不活性化は、受精後の胚珠に必要のない花粉管の誘引を防ぐことで、少ない花粉でも無駄なく種子をつくるために必要と考えられています。今回の研究で、不活性化の仕組みの全貌が明らかとなりました。これを発展させることで、将来、作物の収穫を増やすことができるかもしれません。
植物の細胞は動物とは異なり、堅い細胞壁に覆われています。そのため、植物細胞同士が融合するとは考えられてきませんでした。実際、被子植物の成長過程でみられる例外的な細胞融合として、110年以上も前に卵細胞の受精と中央細胞の受精が報告されて以来、その他の融合は知られていませんでした。今回の現象は、受精に次ぐ新たな細胞融合の発見です。しかも、これまで成長過程でみられる細胞死といえば、動植物を含めて、死ぬべき細胞自身が自殺のプログラムを実行していると考えられてきましたが、今回の現象は細胞融合によって融合相手が不活性化されます。これは植物細胞がもつ静的なイメージを大きく変えるだけではなく、全く新しいタイプの細胞死があることを提示することで、これまでの細胞死の定義を揺るがす可能性をもった発見です。これらの点から、今回の現象は教科書を書き換える程の成果といえます。また、受精以外の細胞融合を引き起こすプログラムの存在が示されたことから、植物における新たな細胞融合技術の開発にもつながることが期待されます。
論文情報:
"Rapid elimination of the persistent synergid through a cell fusion mechanism" by Daisuke Maruyama*, Ronny Völz, Hidenori Takeuchi, Toshiyuki Mori, Tomoko Igawa, Daisuke Kurihara, Tomokazu Kawashima, Minako Ueda, Masaki Itoh, Masaaki Umeda, Shuh-ichi Nishikawa, Rita Groß-Hardt and Tetsuya Higashiyama, Cell (2015)
DOI: 10.1016/j.cell.2015.03.018
リンク:
- プレスリリース
- JST 細胞死をもたらす新しい細胞融合現象の発見 ~受精後の花粉管誘引停止の全貌が明らかに~
- Nagoya University Research



丸山特任助教 東山教授
丸山 大輔特任助教: http://www.itbm.nagoya-u.ac.jp/ja_backup/members/d-maruyama/
東山 哲也教授: http://www.itbm.nagoya-u.ac.jp/ja_backup/members/t-higashiyama/
関連記事・報道:
- NHK 「植物の新しい細胞融合現象発見」 (2015年4月24日)
- 中日新聞 「植物の「細胞融合」で新たな仕組み発見 名大グループ」 (2015年4月24日 朝刊 総合3面)
- 読売新聞 「植物細胞が融合「教科書書き換える発見」...名大」 (2015年4月24日 朝刊 社会30面)
- Yahooニュース 「植物細胞が融合「教科書書き換える発見」...名大」 (2015年4月24日)
- マイナビニュース 「名大、新しい細胞融合現象を発見 - 「教科書を書き換えるほどの成果」」 (2015年4月24日)
- 日経産業新聞 「花粉、効率よく受精 名古屋大 精細胞の誘導解明」 (2015年4月24日)
- Alpha Galileo 「細胞死をもたらす新しい細胞融合現象の発見 〜受精後の花粉管誘引停止の全貌が明らかに〜」 (2015年4月27日)
- 日経バイオテク 「「バイオイメージング最前線」第1回」 最先端の生命科学研究に不可欠なイメージング技術日本の強みの最前線を紹介」 (2015年4月27日)
- EurekAlert!日本語 「細胞死をもたらす新しい細胞融合現象の発見」 (2015年4月24日)
- ライフサイエンスレビュー 「植物の受精における細胞融合による残存助細胞の迅速な排除」 (2015年5月7日)
- 日経バイオテク 「「バイオイメージング最前線」第3回」 経時観察でまだまだ見つかる新現象」 (2015年7月30日)
2015-04-24