研究ハイライト
花粉管は核がなくても胚珠に辿り着く ~世界で初めて核を持たない花粉管の作出に成功~
横浜市立大学の丸山大輔 助教、立命館大学の元村一基 助教らの研究チームは、名古屋大学との共同研究により、モデル植物シロイヌナズナを用いることで、世界で初めて細胞質中に"細胞核"が存在しない花粉管を作出することに成功しました。さらにこの細胞核を除いた花粉管(オス)が、核を持つ正常な花粉管と同様に、雌しべの奥にある生殖器官の "胚珠*1(メス)"へ正確に辿り着く能力を保持していることを明らかにしました(図1)。
細胞核は高校生物の教科書の冒頭で紹介される、遺伝子の発現をつかさどる存在です。花粉管でも常に先端にある核が、花粉管の長距離に渡る伸長や正確な伸長方向制御に必要であると考えられてきました。こうした常識を覆し、細胞核からの新規の遺伝子発現を必要とせず胚珠へと方向転換して、胚珠内の卵細胞へ辿り着く能力を保持していることが示されました。
研究成果は、2021年4月22日10時(英国夏時間)、英国Natureグループが発行するオンライン科学誌「Nature Communications」に掲載されました。
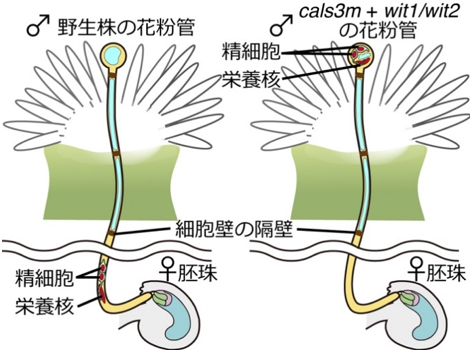
図1 細胞核を持たない花粉管も胚珠を目指す
花粉管の伸長能力を調べるために、培地上に置いた胚珠に花粉管が誘引されるかどうか調べた。
左:野生株の場合、雌しべの切り口から伸びた花粉管は胚珠へ向かって方向を変えて進入する。
右:wit1/wit2変異体でcals3mを発現させた花粉管では、2つ精細胞と栄養核が基部側から動かず細胞壁の隔壁によって閉じ込められた状態になる。しかし花粉管は本来の伸長能力を失わず、野生株と同様胚珠に進入する。
【研究の背景】
動植物の細胞が形を変えるにはその変化に合わせて遺伝子の発現が変動することが重要であると考えられています。したがって花粉管が胚珠へ向かう最中も、その伸長とともに多くの遺伝子の発現が変動することで"花粉管が胚珠まで到達する能力"が制御されていると考えられてきました。
これらの遺伝子発現は"細胞核"の中で行われますが、花粉管には、花粉管細胞の核である"栄養核"と、花粉管中に内包される精細胞の核である2個の"精核"、合計3つの細胞核が含まれています(図2)。これらの細胞核がどのように花粉管の伸長を制御しているのかを明らかにするためには、それぞれの細胞核を欠損させる方法が一般的に考えられます。しかし花粉管の中は膨圧が高く維持されているため、レーザーなどを用いて細胞核を人工的に除去しようとしても花粉管自体が破裂してしまいます。そのため過去に花粉管伸長における細胞核の機能を検証できた例はほとんどありませんでした。
【研究内容】
当初、我々の研究グループでは、精細胞に細胞壁成分が乏しいことに着目し、これが精細胞の正常な機能に重要なのではと考えました。そこで、細胞壁構成成分のひとつであるカロース*2を過剰に合成する能力をもつ変異型酵素の遺伝子cals3m*3を精細胞で発現させて機能解析を試みたところ、興味深いことに精細胞の輸送が阻害され花粉管基部付近に留まることが分かりました(図2)。我々はこの現象が花粉管伸長における細胞核の機能の検証に使えると考え、栄養核の輸送異常を示すwit1/wit2*4変異体でcals3mによる精細胞の輸送異常を誘導したところ、細胞核全てが花粉管基部に残されて先端側に核を持たない花粉管を作出することに成功しました(図3)。
このcals3mを発現するwit1/wit2変異体の花粉管では、伸び始めてから数時間で花粉管内に作られる細胞壁の隔壁が3つの細胞核を花粉管先端から完全に分断します。そのため、先端側に遺伝子発現産物は供給されなくなります(図3)。しかし、驚くべきことにこの状態でもcals3mを発現するwit1/wit2変異体の花粉管は、野生型の花粉管と遜色なく伸長を続け、卵細胞のある胚珠の位置を認識して正確にその中に進入できました(図1)。以上の結果より、花粉管は細胞核に頼らずに胚珠を目指す潜在能力を持つことがわかりました。
本研究でわかった花粉管機能の自立性は、様々な環境条件でも変わらずに子孫を残す花粉管の仕組みの理解に役立つでしょう。また、cals3mを発現させることで精細胞の輸送異常を引き起こせることがわかり、この花粉管を用いて精細胞の輸送メカニズムの解明が進むと期待されます。さらに、精細胞の移動異常をもつ花粉管を受け入れた胚珠は、デンプンに代わり砂糖を貯めるイネの未授精胚珠に似た肥大を示すこともわかり、今後、本研究が様々な植物で砂糖を生産させるために役立つかもしれません。
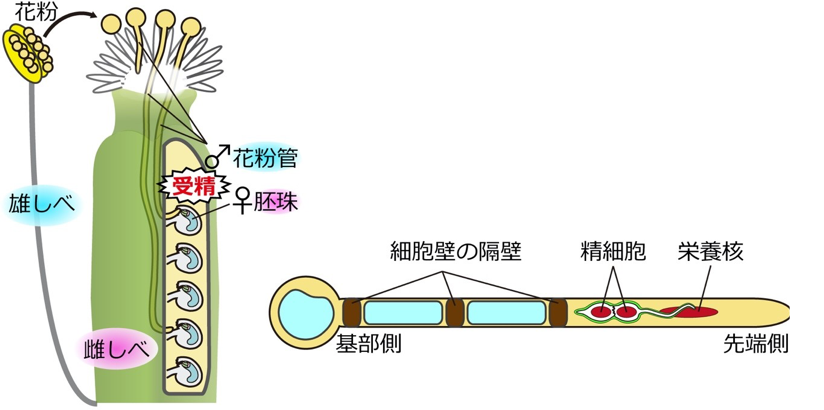
左:花粉が雌しべの先端に受粉すると、細長い花粉管が伸び始める。雌しべの奥深くの胚珠まで伸長すると、花粉管中の精細胞を胚珠内に放出して受精する。
右:花粉管の先端側では、栄養核とそれに連なった2つの精細胞が存在する。一方基部側では、伸長にともなって細胞壁の隔壁が一定間隔で作られ、ほとんどの細胞内構造を、活発に伸長する先端側に維持している。
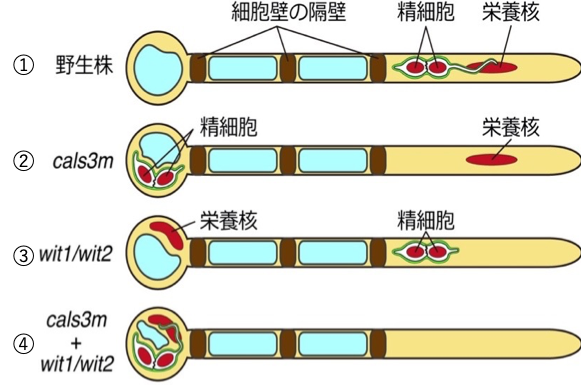
1) 野生株では栄養核と2つの精細胞が花粉管の先端に位置する。
2) cal3mを発現すると2つの精細胞は基部側に留まる。
3) 逆にwit1/wit2変異体では、栄養核が基部側に留まる。
4) さらに、wit1/wit2変異体でcals3mを発現すると、栄養核も精細胞もともに基部側に留まり、先端に細胞核が存在しない花粉管になる。
【用語解説】
1)胚珠:
花粉管を受け入れた後に種子になるメス側の組織。卵細胞や中央細胞といった配偶子を含む複数の細胞からなっている。
2)カロース:
高等植物が持つ多糖の一種。その構造からβ-1,3-グルカンとも呼ばれる。植物細胞の細胞壁構成成分の一種であり、蓄積量を変化させることで細胞壁の硬さや細胞間の分子の流入量を調節する役目を持つ。
3)cals3m:
シロイヌナズナのカロース合成酵素CALLOSE SYNTHASE3 (CALS3)/GLUCAN SYNTHASE-LIKE12 (GSL12)に数塩基の変異を加えることで、過剰にカロースを合成する能力が備わった酵素の遺伝子。cals3mが発現する細胞では過剰量のカロースが堆積する。
4)wit1/wit2:
WPP domain-interacting tail anchored protein 1とWPP domain-interacting tail anchored protein 2の変異体の略称。2種類のタンパク質(WIT1/WIT2)は栄養核膜上に局在し、花粉管内の細胞骨格に沿って物質を運ぶミオシンというモータータンパク質に結合することで、花粉管内における栄養核の輸送に関わっていると考えられている。
【論文情報】
雑誌名:Nature Communications
論文タイトル:Persistent directional growth capability in Arabidopsis thaliana pollen tubes after nuclear elimination from the apex(シロイヌナズナ花粉管は無核の状態でも正常に伸長して胚珠へ到達する能力を保持している)
著者:元村 一基、武内 秀憲、野田口 理孝、土 春菜、竹田 篤史、木下 哲、東山 哲也、丸山 大輔
DOI:https://doi.org/10.1038/s41467-021-22661-8
2021-04-23