研究ハイライト
植物の気孔はなぜ2つの細胞が穴を囲んだ構造になっているのか? ~指令遺伝子ミュートの働きから見えてきた生物の形作りの巧妙な仕組み~
名古屋大学トランスフォーマティブ生命分子研究所(ITbM)の鳥居 啓子主任研究者/客員教授(米国ワシントン大学教授•ハワードヒューズ医学研究所正研究員)、名古屋大学高等研究院Soon-Ki Han(ハン・スンキ)特任助教、九州大学大学院医学研究科の三浦 岳教授、理化学研究所の遠藤 高穂上級研究員らの研究グループは、植物学、バイオインフォマティクス、および数理モデルを組み合わせることにより、陸上植物の気孔の形を作る細胞の分裂が対称的に一回のみ起きるメカニズムを解明しました。
気孔は、スギゴケやシダを含むほぼ全ての陸上植物に存在し、光合成のためのガス交換や水分の制御を担っています。気孔が環境に応じて開閉し、機能するためには、2つ(一対)の孔辺細胞が穴を囲んだ構造が必須であり、実際、どのような植物種でもこの気孔構造がみられます。気孔は、元になる細胞が一回だけ対称に分裂することによってできますが、どのようにして細胞分裂が厳密に一回だけ起こるのかはわかっていませんでした。
はじめに研究グループは、気孔の分化を指令するマスター転写因子ミュート(MUTE)の直接標的遺伝子の網羅的解析を行いました。その結果、MUTEは、気孔の対称分裂に必要な細胞周期因子を誘導すると同時に、これら細胞周期因子を直接抑制する転写因子を直接誘導することにより、気孔の分化過程において細胞分裂が一回だけ厳密に起こるよう統御していることがわかりました。
研究グループは、数理モデルを用いて、この制御ネットワークに異常が生じた場合に起こる気孔の発生異常を予測し、 実験で検証しました。このMUTEによる統御様式はフィードフォワード回路と呼ばれ、動物や酵母、大腸菌などの迅速な遺伝子発現制御に広くみられます。そのため、本研究は、動植物、原核•真核生物を問わず形作りの基盤原理に迫るものであり、気孔の発生制御を介して作物の改良にも貢献が期待されます。
この研究成果は、平成30年 5月7日正午(米国東海岸夏時間)米国科学雑誌Developmental Cell に掲載されました。
研究の内容:
【ポイント】
■ モデル植物シロイヌナズナの気孔の分化過程における全遺伝子発現変動の挙動を解析しました。
■ 気孔の分化を指令する転写因子ミュート(MUTE)は、気孔系譜を作る姉妹転写因子スピーチレス(SPEECHLESS)の標的遺伝子を引き継いで、その発現パターンを強制的に変えることにより、気孔系譜の幹細胞を分化させることがわかりました。
■ 転写因子ミュート(MUTE)は、新奇細胞周期因子(CYCD5;1)を始めとする一連の細胞周期因子を直接誘導するとともに、これら細胞周期因子を直接抑制する転写因子(ミュートの姉妹因子であるファーマ(FAMA)、およびフォーリップス(FOUR LIPS(FLP))を直接誘導することがわかりました。
■ ミュート(MUTE)を頂点とした細胞周期因子の発現制御は、1型インコヒーレント・フィードフォワード回路と呼ばれるもので、この遺伝子制御ネットワーク回路によって、気孔の分化過程に細胞分裂が厳密に一回だけ起こり、2つの孔辺細胞が穴を囲んだ形状の気孔ができることが明らかになりました。
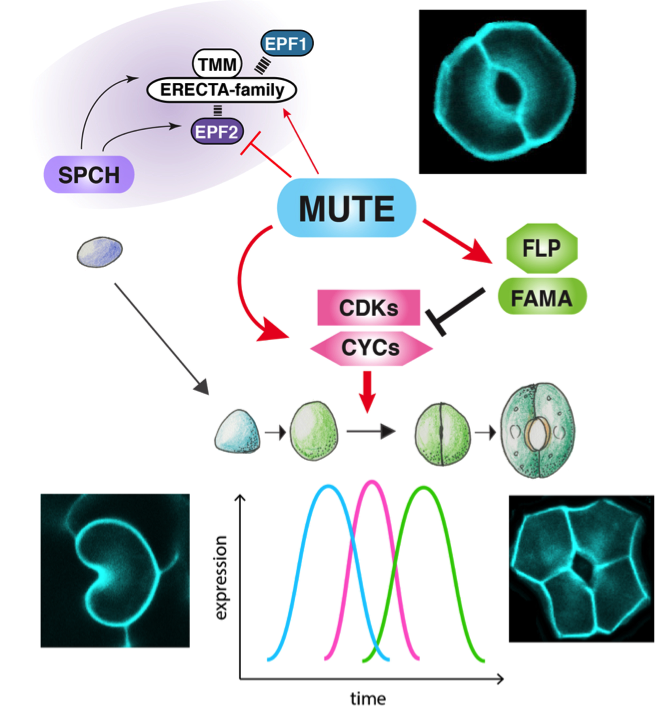
図1 MUTEによる気孔の分化と細胞分裂の回数の制御ネットワーク(概念図)
この制御回路をつなぎかえると、正常な2つの孔辺細胞が穴を囲んだ形状の気孔(右上)ではなく、1つの孔辺細胞だけの気孔(左下)や、逆に4つの孔辺細胞からなる気孔(右下)ができる
【研究背景と内容】
子供の頃、理科の実験で、植物の葉の表皮の気孔を観察したことがある人もいるのではないでしょうか。顕微鏡で見る気孔は、2つの(一対の)唇型の孔辺細胞がミクロサイズの穴を囲んだ形をしており、表皮に点在する気孔は少し不思議で不気味かもしれません。植物は、気孔を介して光合成に必要な二酸化炭素を取り込みます。また、気孔から蒸散する水蒸気は、私たちが汗をかくように葉の表面温度を下げる効果を持ちます。一方、乾燥条件下では、気孔が閉まることにより枯死を防いでします。そのため、どの植物種でも気孔は2つの孔辺細胞が穴を囲んだ形をしています。一体どうして、種を超えて安定した気孔の形状ができるのでしょうか。2つの孔辺細胞は、元になる細胞である孔辺母細胞が一回だけ対称に細胞分裂することによってできます。しかし、気孔の発生過程で対称分裂の回数がなぜきちんと制御されているのかわかっていませんでした。
研究グループらは、過去に気孔の発生は、3つの姉妹転写因子であるSPEECHLESS (SPCH), MUTE, FAMAが順々に作用することにより起こることを発見しました。これら3つの転写因子のうち、MUTEを強制的に発現させると、植物表皮の全てが気孔になることがわかっています(図2)。そこで、MUTEは、気孔を作る遺伝子発現プログムを作動させると仮説を立て、MUTEによって制御される遺伝子を網羅的に同定しました。
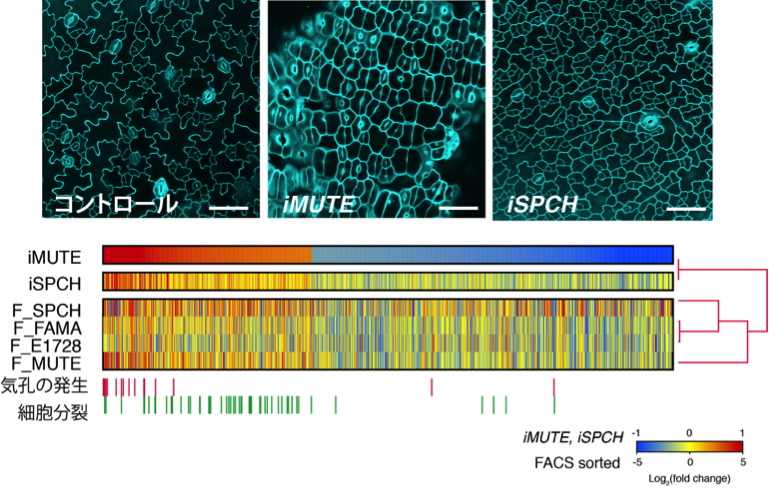 図2 iMUTEにより制御される遺伝子群の同定
図2 iMUTEにより制御される遺伝子群の同定
上:シロイヌナズナ芽生え子葉の顕微鏡写真(左からコントロール;iMUTE-表皮全てが気孔になる;iSPCH-表皮全てが気孔系譜の幹細胞になる)。下: iMUTEによって発現が誘導(赤)抑制(青)される全遺伝子のヒートマップと既存のデータとの比較。iMUTEで発現誘導される遺伝の多くは、気孔の発生および細胞分裂に関わるものであった。
その結果、MUTEが誘導する遺伝子の多くは、気孔の発生・分化に関わるもの、そして細胞分裂・細胞周期に関わるものであることがわかりました。気孔の発生・分化に関わる標的遺伝子の解析から、MUTEは、SPCHの標的遺伝子を受け継ぐと同時に、気孔の幹細胞状態に関わる特定のシグナル因子を抑制することにより、気孔系譜の細胞を未分化な幹細胞状態から分化状態へとスイッチさせることがわかりました。
次に、MUTEが誘導する一群の細胞分裂・細胞周期に関わる因子に着目しました。それらの中には、新奇のD型サイクリン(細胞分裂を引き起こす因子)であるCyclinD5;1が含まれていました。D型サイクリンは、我々ヒトを含む動物や植物において細胞分裂を誘起することが知られています。実際に、CyclinD5;1を発生停止したmute変異体の気孔系譜の幹細胞に導入すると、対称分裂様の異常分裂を起こすことが確認できました。さらに、MUTEは、姉妹転写因子FAMAと、別の種類の転写因子FOUR LIPS (FLP)の発現を直接誘導することがわかりました。FAMAとFLPは、どちらも孔辺母細胞の分裂の抑制作用を持つことが報告されています。
これらの知見から、MUTEは、細胞周期因子と、細胞周期因子を抑制する転写因子の両者を誘導することがわかりました。因子X(ここではMUTE)が、因子Y(ここではFAMA/FLP)と因子Z(ここでは細胞周期遺伝子)を誘導する一方、因子Yは因子Zを抑制する、という制御回路は、1型インコヒーレント・フィードフォワード回路と呼ばれており、因子Zの素早いピークを一回だけ作り出すことができる回路として知られています(図3)。そこで、研究グループは、数理モデルと摂動実験を組み合わせることにより、MUTEが気孔母細胞の細胞分裂を一回だけ起こすよう統御する可能性を検証しました。
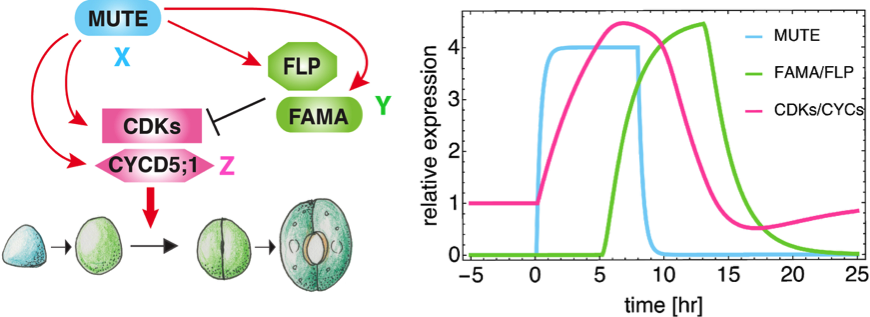 図3 1型インコヒーレント・フィードフォワード回路(左)による細胞周期因子の発現ピークのシミュレーション(右)
図3 1型インコヒーレント・フィードフォワード回路(左)による細胞周期因子の発現ピークのシミュレーション(右)
このフィードフォワード回路の摂動実験として、まず、XとYが同時にONになってしまう場合を予測しました。その場合、シミュレーションでは、細胞周期因子の発現が低く抑えられます。実際に、そのようなシロイヌナズナ芽生えを作出したところ、対称分裂が起きず、一つの孔辺細胞からなるゼリービーンズのような形の気孔ができました。この気孔には穴がないため、もちろん気孔としての働きをすることはできません(図4)。
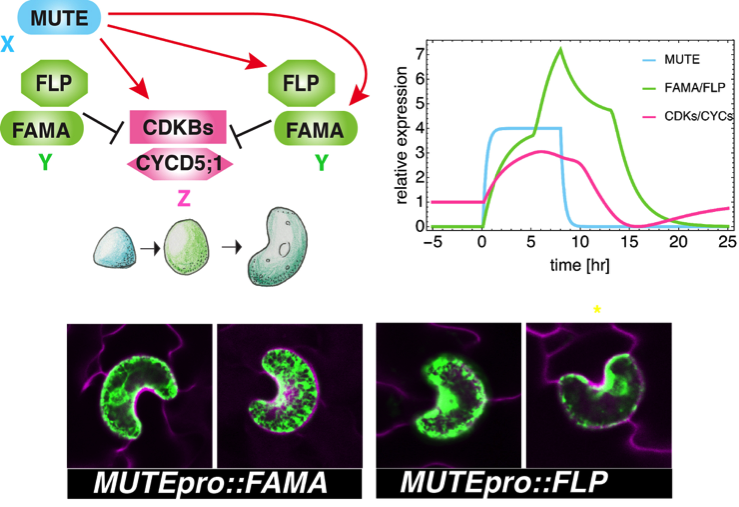 図4 MUTEを頂点とするフィードフォワード回路の摂動実験1:細胞周期因子を抑制するFAMAとFLPの早すぎる発現により、1つの孔辺細胞のみからなるゼリービーンズのような形の気孔ができる。模式図(上左)、細胞周期因子の発現ピークのシミュレーション(上右)、実験による検証(注:このゼリービーンズ様の気孔は、孔辺細胞マーカーGFPを発現しているため、緑色の蛍光を発しています)
図4 MUTEを頂点とするフィードフォワード回路の摂動実験1:細胞周期因子を抑制するFAMAとFLPの早すぎる発現により、1つの孔辺細胞のみからなるゼリービーンズのような形の気孔ができる。模式図(上左)、細胞周期因子の発現ピークのシミュレーション(上右)、実験による検証(注:このゼリービーンズ様の気孔は、孔辺細胞マーカーGFPを発現しているため、緑色の蛍光を発しています)
次に、逆に早すぎる細胞周期因子の発現は、気孔の形成にどう影響するか調べました。FAMA転写因子による抑制が強いため、famaを欠損する背景をシミュレーションと摂動実験に用いています。予測どおり、細胞の過剰な対称分裂が起こることがわかりました。
この1型インコヒーレント・フィードフォワード回路は、入力シグナル(すなわちMUTEの発現)が一定で継続的にONになったとしても、細胞周期遺伝子の発現ピークパターンを一回だけ作り出すことができます。そのため、MUTEを強制的に過剰発現させたとしても対称分裂は一回だけ起こり 、その結果、ほとんどの気孔は、2つの孔辺細胞が穴を囲んだ形になることも説明できました(図2:iMUTEの顕微鏡写真参照)。しかし、数理モデルから、MUTE過剰発現下では細胞分裂が全く生じない場合と過剰な細胞分裂が生じる場合の両方の変異が起こり得ることが予測できました。実際に詳細に観察した結果、この理論予測が裏付けられました。
今回解析したSPCH, MUTE, FAMA転写因子は、気孔を持つ全ての陸上植物で気孔の発生を制御していると考えられます*。そのため、今回発見したMUTEによる、気孔の対称分裂が一回だけ起こる制御メカニズムは、陸上植物に普遍的であると想定されます。また、1型インコヒーレント・フィードフォワード回路は、大腸菌や酵母などの遺伝子発現回路として広く使われているモチーフです。そのため、本研究は、動植物、原核•真核生物を問わず形作りの基盤原理に迫るものです。また、本回路を繋ぎかえることにより気孔の孔辺細胞の数を作り変えることができるため、そのような系を用いて、進化の過程で洗練された気孔の形状と生理機能を統合的に調べることも可能となります。
*注:スギゴケなど初期陸上植物では、SPCH, MUTE, FAMAは3つの姉妹転写因子に進化しておらず、祖先型ハイブリッド転写因子として機能しています。
論文情報:
This article "MUTE Directly Orchestrates Cell-State Switch and the Single Symmetric Division to Create Stomata" by Soon-Ki Han, Xingyun Qi, Kei Sugihara, Jonathan H. Dang, Takaho A. Endo, Kristen L. Miller, Eun-Deok Kim, Takashi Miura, Keiko U. Torii was published online in Developmental Cell.
DOI: 10.1016/j.devcel.2018.04.010
リンク:


関連記事・報道:
- 九州大学 「なぜ気孔は1対の細胞で囲まれる?~植物の形作りの謎に数学で迫る~ 」(2018.5.9)
- 日本経済新聞 「九大、植物の気孔を形作る細胞がただ1回だけ正しく分裂してできるメカニズムを解明」(2018.5.9)
- 日本の研究.com 「植物の気孔はなぜ2つの細胞が穴を囲んだ構造になっているのか?」(2018.5.10)
- 科学新聞「植物の気孔形成の仕組み解明 細胞分裂「左右対称に一回だけ」」(2018.5.25
2018-05-08