研究ハイライト
「のり」が「はさみ」を連れてくる 〜植物細胞のユニークな「微小管」形成の仕組みを解明〜
国立大学法人東海国立大学機構 名古屋大学トランスフォーマティブ生命分子研究所(WPI-ITbM※)の八木 慎宜 博士、中村 匡良 特任講師らは、奈良先端科学技術大学院大学の橋本 隆 教授、カーネギー研究所(米国)のデイビッド エアハルト 博士、東京大学の松永 幸大 教授らとの共同研究で、動物や酵母では「微小管」を「微小管形成中心」注1)につなぎ留める「複合体」が、「微小管形成中心」を持たない植物では独自の機能を持つことを明らかにしました。
細胞骨格の一つである「微小管」は、細胞内でポリマー状の構造をしており、その配向パターンを変化させることで、染色体の分離や細胞極性の制御、細胞の形態形成など、生物の生存に不可欠な活動に寄与しています。植物では、高度に組織化された「表層微小管」注2)の配向パターンが、成長過程や環境シグナルに応じてダイナミックに変化し、変動する細胞内外の環境に適応しています。植物の「表層微小管」のパターン形成には、「微小管」の形成と切断が重要であると考えられています。特に、「微小管形成開始点部位」を「微小管」切断タンパク質「カタニン(日本刀から命名)」が特異的かつ効率的に切断することにより、新たに形成された「娘微小管」を細胞表層に遊離し、「微小管」同士の相互作用を促すことが、「微小管」のパターン形成に必須です。しかしながら、「微小管形成部位」に「カタニン」がリクルートされる分子機構は長らく不明でした。
本研究により、「新規微小管」をその生成部位にとどめておく『のり』分子が、「新規微小管」を基部で切断する『はさみ』分子を連れてくるという、相反する作用を持つというユニークな生命現象が明らかになりました。この2つの機能により、「微小管」切断が成長過程や環境シグナルに応じて高度に制御可能になります。この成果は、植物の形作りを制御する「微小管」の配向パターンの形成メカニズムの解明に繋がる重要な発見です。今後、この知見を発展させ、細胞骨格ネットワークを人為的に制御することで、細胞形態を改変し、環境応答効率を向上させる技術の開発が期待されます。
本研究成果は、2021年6月17日18時(日本時間)付英国科学誌「Nature Communications」に掲載されました。
【ポイント】
- 動植物で保存されている細胞因子が、植物細胞では保存機能に加えて新たな機能を獲得している。
- 「微小管」を細胞内特定箇所につなぎ留める「のり」分子が、植物細胞ではその箇所を切断する「はさみ」分子を連れてくる。
- 「のり」と「はさみ」の共同作業により細胞内環境に適応した微小管パターンの形成が可能となる。
【研究の背景】
「微小管」は、動物細胞で見られる中心体を中心とした放射状配列や、異方性に伸長する植物細胞の中心体を持たない細胞で見られる横方向の「表層微小管」など、様々なパターンを形成しています。中心体のような「微小管形成中心」から新たに形成された「微小管」は、形成中心に固定され、そこから切り離されます。この固定と切り離しの制御が「微小管」の配向パターンに寄与していると考えられますが、詳細なメカニズムは分かっていません。
植物細胞では、「表層微小管」上に結合した「γチューブリン環状複合体」注3)から「微小管」は形成され、新規に形成された「微小管」は「微小管」切断因子「カタニン」の機能により切り離されます(図1)。「カタニン」は「微小管形成中心」と「微小管交差部位」に集積され「微小管」を切断しますが、その集積メカニズムは全くわかっていませんでした。
【本研究の成果】
本研究グループは、酵母や動物細胞で「微小管」を「微小管形成中心」につなぎ留める「Msd1-Wdr8複合体ホモログ」注4)が植物に存在することを明らかにしました。遺伝学的手法とライブセルイメージングを組み合わせることで、シロイヌナズナの「Msd1-Wdr8」複合体が細胞表層に散在する「微小管形成部位」に局在し、「表層微小管形成部位」を安定化することを突き止めました(図1)。
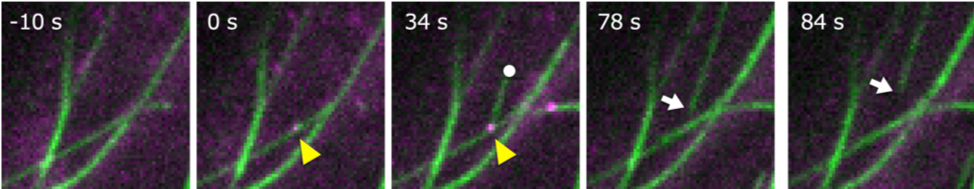
図1:植物表層微小管形成過程の「微小管」と「Wdr8」のライブセルイメージング
「微小管」を緑色の線でWdr8をピンクの点で示し、「Wdr8」が「微小管」上に現れた場面を0秒(0 s)としている。「Wdr8」が集積する「微小管形成部位」(黄色い矢尻)から新規に形成された「微小管」のプラス端を○印で示している。「微小管」は「カタニン」により形成部位から切り離される(矢印は「微小管」のマイナス端)。
さらに、「新規微小管」を切り離す因子である「カタニン」を「微小管形成部位」に集積するために利用されていることが明らかとなりました。「微小管交差部位」には、「Msd1-Wdr8複合体」は局在せず、「微小管形成部位」特異的に「カタニン」の集積を制御していました(図2)。
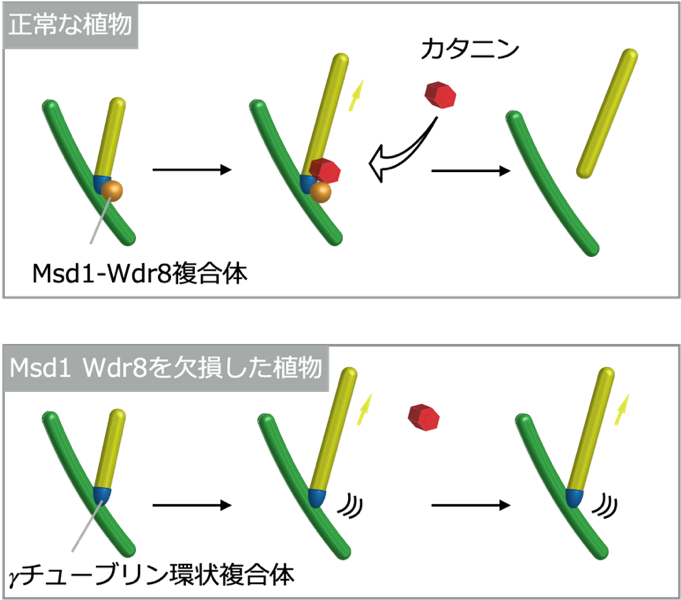
図2:正常な植物と「Msd1」「Wdr8」を欠損した植物体における微小管形成の様子
既存の「微小管」上に結合した「γチューブリン環状複合体」から「微小管」は形成される。「微小管形成部位」に「Msd1-Wdr8」が結合すると、「カタニン」が「微小管形成部位」にリクルートされ、「微小管の形成部位」からの切り離しが促される。「Msd1」もしくは「Wdr8」を欠損した植物では、形成部位が安定せず、「カタニン」が「微小管形成部位」に集積できず、「微小管」の切り離しが起こらない。
本研究により、植物のユニークな「微小管形成」の仕組みの一端が明らかとなりました。「Msd1-Wdr8複合体」は、動物や酵母にも存在するタンパク質です。動物や酵母で見られる「微小管形成中心」に「微小管」をつなぎ留める機能に加え、「Msd1-Wdr8複合体」は「カタニン」の局在を制御することで、「微小管」の切り離しをより厳密な制御下に置く能力を植物に与えていると考えられました。「Msd1-Wdr8」の働きをより詳細に調べることで、植物における「微小管形成」のメカニズムとその進化の過程が明らかになることが期待されます。
【用語解説】
注1)微小管形成中心:
動物細胞の中心体のように、微小管を形成する中心となる細胞小器官。
注2)表層微小管:
植物間期細胞の細胞膜内側にネットワーク状に存在する微小管構造。細胞壁成分セルロース繊維の沈着を介して植物細胞のかたちをつくり上げる。
注3)γチューブリン環状複合体:
微小管の重合核となるγチューブリンを含む複合体。微小管の鋳型となるリング状構造をしている。
注4)ホモログ:
異なる生物種で類似性の高い塩基配列を持つ遺伝子のこと。共通の遺伝子から由来している可能性が高い。
【論文情報】
雑誌名:Nature Communications
論文タイトル:An anchoring complex recruits katanin for microtubule severing at the plant cortical nucleation sites(微小管切断因子カタニンを表層微小管形成部位にリクルートする微小管安定化複合体の発見)
著者:Noriyoshi Yagi, Takehide Kato, Sachihiro Matsunaga, David W. Ehrhardt, Masayoshi Nakamura, Takashi Hashimoto(八木 慎宜、加藤 壮英、松永 幸大、デイビッド W. エアハルト、中村 匡良、橋本 隆)
DOI: 10.1038/s41467-021-24067-y
2021-06-19