研究ハイライト
花粉管を誘引する際の鍵と鍵穴を解明 ~植物の受精において重要な花粉管誘引物質と受容体の結合時の構造が明らかに~
名古屋大学トランスフォーマティブ生命分子研究所(ITbM)の東山 哲也教授/副拠点長、大学院理学研究科の長江 拓也大学院生、武内 秀憲特任助教(オーストリア・グレゴールメンデル研究所 日本学術振興会海外特別研究員)らの研究グループは、中国・精華大学のJijie Chai教授らとの共同研究で、花粉管誘引ペプチドである「ルアー」と花粉管に存在するルアーの受容体である「PRK6」タンパク質が結合した状態での共結晶構造解析に初めて成功しました。これにより、ルアーとその受容体が、鍵と鍵穴が合うように正確に結合する仕組みが明らかとなりました。
精細胞を運ぶ花粉管が、迷わずに卵にたどり着くために起こる花粉管の誘引は、植物の受精において重要な過程です。今回の発見により、同種の植物間でのみ、ルアーが正確に働く仕組みの解明や、これまで難しかった異種間の交雑による有用品種作出への応用が期待できます。
この研究成果は、英国科学雑誌Nature Communications電子版に掲載されました。
【ポイント】
■ 花粉管誘引ペプチドである「ルアー」と花粉管に存在する「PRK6」タンパク質が結合する仕組みを解明した。
■ ルアーに対してPRK6が真の受容体であることが証明された。
■ ルアーとPRK6の結合および花粉管誘引に重要なアミノ酸が明らかとなった。
研究の内容:
【研究背景と内容】
被子植物の受精では、花粉から伸びだした花粉管が、正確に卵のある場所まで到達することで受精が起こります。花粉管は、「花粉管ガイダンス(誘引)」と呼ばれる多段階の伸長方向制御を受けることで、道に迷わず卵まで到達します。特に、最終段階においては、卵のとなりに2つある助細胞という特殊な細胞から分泌されるルアーと呼ばれる誘引物質が分泌され、同種の花粉管が正確に誘引されます(Higashiyama et al., 2001, Science; Okuda, Tsutsui et al., 2009, Nature)。モデル植物であるシロイヌナズナにおいてもルアーが発見され(Takeuchi and Higashiyama, 2012, PLoS Biology)、そのルアー(LURE1)に対する受容体として、2つの研究グループから、それぞれMDIS-MIK注1) (Wang et al., 2016, Nature)と、PRK6注2)(Takeuchi and Higashiyama, 2016, Nature)という異なるタンパク質が報告されていました。受容体に結合する分子をリガンドと言います。リガンドと受容体の関係は、鍵と鍵穴にたとえることができます。ルアー(リガンド)がどの受容体にどのように結合するのかは、これまで明らかではありませんでした。
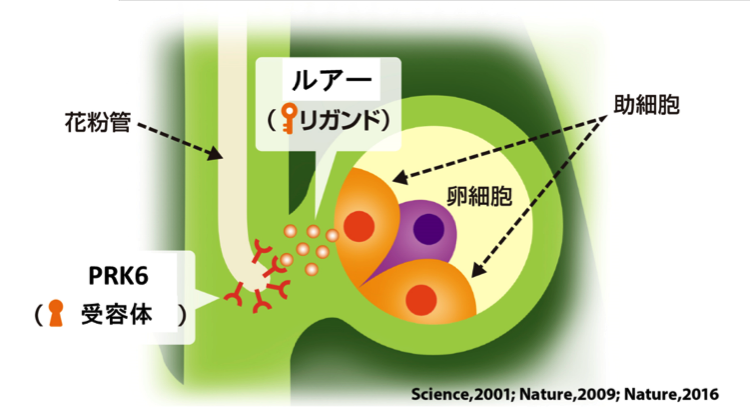 図1.雌しべの中でルアーによって誘引される花粉管。新学術領域研究「植物新種誕生原理」ホームページより、改変のうえ転載。
図1.雌しべの中でルアーによって誘引される花粉管。新学術領域研究「植物新種誕生原理」ホームページより、改変のうえ転載。
本研究では、ルアーがどのように受容体に結合するのかを解明するため、X線結晶構造解析に着目しました。はじめに、昆虫の培養細胞系を用いて、シロイヌナズナのルアー(LURE1.2)や受容体タンパク質(細胞外領域のみ)をつくり、ルアーと結合するタンパク質を調べました。その結果、ルアーはPRK6と特異的に結合することが示されました。同じPRKファミリーのPRK3、PRK4、PRK5や、他グループから報告されていたMIK1、MIK2タンパク質とは結合しませんでした。
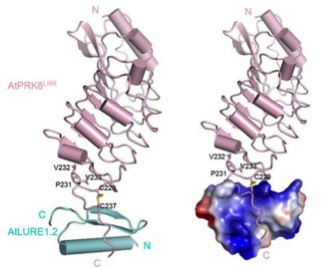 図2.ルアー (AtLURE1.2) と受容体PRK6 (AtPRK6)の共結晶構造。右ではルアー表面の電荷を示している(青が正の電荷)。今回の論文 (Zhang, Liu, Nagae et al., 2017, Nature Communications) から転載。
図2.ルアー (AtLURE1.2) と受容体PRK6 (AtPRK6)の共結晶構造。右ではルアー表面の電荷を示している(青が正の電荷)。今回の論文 (Zhang, Liu, Nagae et al., 2017, Nature Communications) から転載。
そこで、次にルアーとPRK6が結合した状態での結晶構造、すなわち、共結晶構造をX線結晶構造解析の手法で明らかにすることを目指しました。その結果、初めて共結晶構造を解くことに成功しました(図2)。解析の結果、PRK6のロイシンリッチリピートと呼ばれるアミノ酸配列の領域と、膜貫通領域注3)という細胞膜を貫通する領域の間にルアーがはまり込むように結合していることがわかりました。ルアーは細胞膜に近い部分でPRK6と結合していることになります。このルアーが結合している場所をループ領域と呼びます。ルアーの正電荷とPRK6のループ領域の負電荷が、静電的な相互作用に重要でした。また、ルアーが結合したPRK6のループ領域には、ルアーが結合していない場合には見られない分子内ジスルフィド結合注4)という架橋(C229-C237)が見られることもわかりました。この架橋によるループ領域の安定化が、ルアーとの結合に重要であることが考えられます。
ルアーとPRK6の結合の仕方には、ユニークな点が2つありました。1つ目は、結合の場所です。これまでに報告されている植物の受容体とペプチドの結合は、普通はロイシンリッチリピートの部分(筒状に描かれた構造が繰り返している図2の上側の領域)で起こります。2つ目の点は、1つのルアーと1つのPRK6が結合し、予想されていたルアーによるPRK6同士またはPRK3との複合体形成の促進は見られなかったことです。一般に、受容体の細胞外領域にリガンドの分子が結合すると、複合体が作られることで細胞の中にシグナルを伝えるケースが多く見られます。今回観察されたルアーとPRK6のユニークな結合は、ルアーがPRK6を介して花粉管の伸長方向を正確にコントロールする仕組みを反映していることがわかりました。
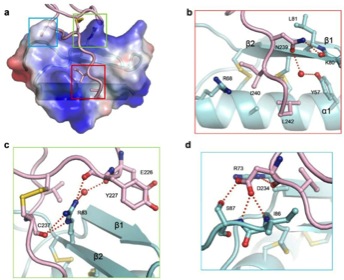 図3.ルアーとPRK6の結合部位の拡大図(aで色別に囲んでいる部分をbからdで拡大している)今回の論文 (Zhang, Liu, Nagae et al., 2017, Nature Communications) から転載
図3.ルアーとPRK6の結合部位の拡大図(aで色別に囲んでいる部分をbからdで拡大している)今回の論文 (Zhang, Liu, Nagae et al., 2017, Nature Communications) から転載
ルアーとPRK6ループ領域の結合をさらに拡大すると、結合に重要なアミノ酸が見えてきます(図3)。このように、結晶構造解析から結合に重要と推定されるアミノ酸を他のアミノ酸に変えて結合性や花粉管の誘引率を調べました(図4)。その結果、ルアー側で83番目のアルギニン(R83)というアミノ酸と、PRK6側で234番目のアスパラギン酸(D234)など、結合にも誘引にも重要なアミノ酸が明らかとなりました。
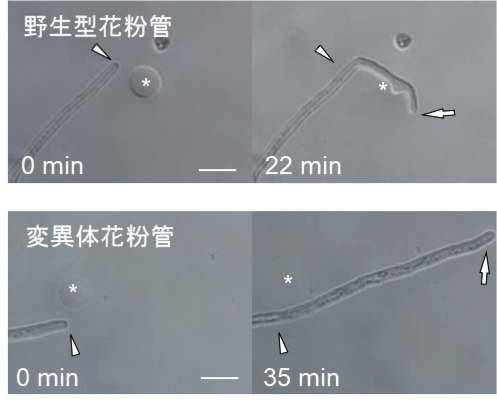 図4.ルアーとPRK6の結合および花粉管誘引に重要なアミノ酸の解析。野生型の花粉管(上)に比べて、ルアーとの結合に重要なPRK6のアミノ酸を変異させた花粉管(下)ではルアーへの誘引率が低下。アスタリスクの部分にルアーを置いて花粉管が誘引されるか観測(スケールバー20㎛)。今回の論文 (Zhang, Liu, Nagae et al., 2017, Nature Communications) から改変のうえ転載
図4.ルアーとPRK6の結合および花粉管誘引に重要なアミノ酸の解析。野生型の花粉管(上)に比べて、ルアーとの結合に重要なPRK6のアミノ酸を変異させた花粉管(下)ではルアーへの誘引率が低下。アスタリスクの部分にルアーを置いて花粉管が誘引されるか観測(スケールバー20㎛)。今回の論文 (Zhang, Liu, Nagae et al., 2017, Nature Communications) から改変のうえ転載
【成果の意義】
本研究により、共結晶構造が解かれた誘引物質ルアーとその受容体PRK6は、いずれも名古屋大学の東山哲也教授の研究室によって同定されてきました。それらの共結晶構造が明らかとなり、これらが誘引物質と真の受容体であると証明されたことは、東山教授の研究室が、本研究分野の進展に大きく貢献してきたことを示すものです。
ルアーは、受容体のPRK6を介して、卵がある方向に花粉管の伸長方向を変えますが、なぜ、ルアーがそのように花粉管の伸長方向を精密にコントロールできるのか、未だ、大きな謎が残っています。ルアーがPRK6のループ領域に結合し、さらなる複合体形成を引き起こさなかったことは、ルアーによる花粉管伸長方向制御の仕組みを理解するうえで、極めて重要な知見です。例えば、ルアーを与えるとPRK6がルアー側に集まることが示されていますが(Takeuchi and Higashiyama, 2016, Nature)、ルアーがループ領域に結合することで、PRK6の花粉管上での動きが変化する可能性が考えられます。今後、ルアーおよびPRK6の分子1つ1つの動きを、花粉管表面でリアルタイムに観察することで、謎の解明が進むと期待されます。
また、ルアーとPRK6は、鍵と鍵穴のように、精密に種の認証を行う分子です。それらの結合の仕方が原子レベルで明らかになったことで、鍵と鍵穴が、同種間では効率よく働き、異種間ではうまく働かない理由が明らかになることも見込まれます。さらに、異種間でも効率よく働くように、鍵と鍵穴をデザインすることで、これまで難しかった異種間の交雑が可能になるとことも大いに期待されます。異種間の交雑で誕生してきた新種植物には、世界三大の穀物のパンコムギ、オイルを多量にとることができる西洋ナタネ、また、木綿を得るためのワタなど、多くの有用作物があるため、本研究は、デザインされた交雑育種に向けた非常に重要な研究としても位置付けられます。
【用語説明】
注1) MDIS-MIK:MALE DISCOVERER (MDIS)、MDIS1-INTERACTING RECEPTOR LIKE KINASE (MIK)
シロイヌナズナのルアー(LURE1)に対する受容体として、Wangらによって報告(Wang et al., Nature, 2016)
注2) PRK6:POLLEN-SPECIFIC RECEPTOR-LIKE KINASE 6
シロイヌナズナのルアー(LURE1)の受容に必須な受容体として、武内および東山によって報告(Takeuchi and Higashiyama, Nature, 2016)。PRK6が属するPRKファミリーは、花粉管の発芽や伸長に重要な受容体キナーゼとして有名
注3) 膜貫通領域:生体膜(脂質二重層)に付着する膜タンパク質において、脂質二重層を貫通している領域(疎水性アミノ酸が多く連なる)。ここでは、細胞膜の外側(ルアーを受け取る細胞外領域)と内側(キナーゼ領域などをもつ細胞質領域)をつなぐ領域
注4) 分子内ジスルフィド結合:1つのタンパク質に含まれる2つのシステインの間で作られる架橋構造。ジスルフィド結合はシステインのチオール基(SH)同士が架橋して作られることから、S-S結合とも呼ばれる。
論文情報:
This article "Structural basis for receptor recognition of pollen tube attraction peptides" by Xiaoxiao Zhang, Weijia Liu, Takuya T. Nagae, Hidenori Takeuchi, Heqiao Zhang, Zhifu Han, Tetsuya Higashiyama & Jijie Chai is published online in Nature Communications
DOI: 10.1038/s41467-017-01323-8
リンク:
- プレスリリース
- 中日新聞「植物受精 タンパク質「認証性」 めしべ内仕組み 名大グループ解明」(2017.11.11 朝刊 32面)
- 日刊工業新聞「名大 植物受精の仕組み解明 花粉管誘引物質 たんぱく質と結合」(2017.11.22 朝刊 31面)
- マイナビニュース 「SSHの生徒たちが感じた、科学の面白さとは - 平成29年度スーパーサイエンスハイスクール生徒研究発表会」(2017.08.23)
- AlphaGalileo JP 「花粉管を誘引する際の鍵と鍵穴を解明」(2017.12.29)
- EurekAlert! JP 「花粉管を誘引する際の鍵と鍵穴を解明」(2017.12.29)


関連記事・報道:
2017-12-29