水陸両生植物の気孔の謎 〜水没しても生き延びる仕組みを解明!

【ポイント】
・水陸両生植物Rorippa aquaticaは、水没により逃げ場を失った植物ホルモン「エチレン」が体内に蓄積されることで環境の変化を機敏に感知することがわかった。
・生理学的解析および次世代シークエンサーを使った遺伝子発現解析により、水没時の非常に迅速な「気中葉から水中葉への運命転換の仕組み」を解明した。特に、Rorippa aquaticaでは水中生活につながる進化の過程において、陸上で生育する植物が持つエチレン応答経路や光応答経路がつなぎかわり、水没時に葉の表皮の気孔の形成に関わる遺伝子の発現を抑制する仕組みを獲得したことを明らかにした。
・植物に備わっている環境の変化に機敏に応答する仕組みを知ることは、気候変動から植物を護る方法や、環境が変化しても生産性が落ちない農作物の実現を考えるうえで有用な手がかりになる。
【研究概要】
京都産業大学の池松朱夏博士研究員、木村成介教授、名古屋大学の鳥居啓子主任研究者・客員教授らの共同研究グループは、水陸両生植物Rorippa aquatica(ロリッパ・アクアティカ)を用いて、植物ホルモン・エチレンの働きにより、水没という環境変化が機敏に感知され、葉の気孔の形成にかかわる遺伝子の発現が抑制されるという「気中葉から水中葉への運命転換の仕組み」を突き止めました。
本研究成果は、植物に備わっている能力を用いることにより、環境変動下でも継続的に生産が可能な農業の実現につながることが期待されます。Cell プレス社が発行する国際学術雑誌Current Biology(2023年1月25日付(日本時間))に掲載されました。
【研究体制】
京都産業大学、名古屋大学、テキサス大学(米国)、中国科学院水生生物研究所(中国)、マラーゲ大学(イラン)の共同研究グループ
【研究背景】
今から約4億5千万年前、水中藻類の一部が分岐し、陸地に進出し始めました。これが現在、陸地に繁栄している陸上植物の起源です。陸上植物は、その進化の過程で、陸上の乾燥に耐えるため、「クチクラ」と呼ばれる脂質ポリマーで自らの体を囲うようになり、また、光合成に必要な二酸化炭素を取り込むために、葉に「気孔」と呼ばれる穴を開けるようになりました。したがって、陸上植物を水没させると、気孔が塞がれ、空気の出し入れ(ガス交換)が出来なくなるため枯れてしまいます。
一方、陸上植物が再び水に進出したグループがいます。それが水草です。多くの水草は、厚いクチクラ層を持たず、気孔も作りませんが、表皮からの拡散によってガス交換をすることができます。これによって水草は、水中でも光合成ができるのです。
さて、植物種の中には、陸上でも水中でも生育できるグループがいます。これを「水陸両生植物(amphibious plant)※1」と呼びます。水陸両生植物は河川や湖沼などの水辺に生育し、水没すると水草のように、また干上がると再び陸上植物のように生育することができます。アブラナ科植物の一種である「Rorippa aquatica ※2(以下R. aquaticaと略称)」は北米大陸原産の水陸両生植物で、水没すると葉身を細くし、気孔発生を抑制します。そうすることで、水中生活に適した葉を発生させるのです(図1)。しかし、水陸両生植物が水没という環境変化を感知して、どのように気孔発生を抑制しているのかは、これまでわかっていませんでした。
【研究成果】
【水没に応答した気孔発生の抑制の観察】
気中(陸上)で生育させたR. aquaticaは、気孔を持った葉(気中葉)を作ります。一方、水中で生育させたR. aquaticaの葉を観察すると、気孔がありませんでした(図1)。 そこで我々は、気中で育てたR. aquaticaを水没させて、葉がどのように変化するかを経時的に観察してみました。その結果、成長中の若い葉のでは気孔の発生が抑制され、気中葉から水中葉へ運命転換が行われていること、また、この「気孔発生の抑制」は水没直後におこっており、4日後には葉の表皮における気孔の密度が明らかに低下していることがわかりました。
通常、一度葉にできた気孔を無くすことはできません。R. aquaticaにおいても、気中葉として完成している成熟した葉は、気孔を減らして水中葉になることはできません。しかし、未成熟な成長中の葉は、水没させればすぐに水中葉へ変化できるのです。そして、この素早い変化により、R. aquaticaは、水没しても問題なく成長を継続することができるのです(図2)。
【気孔発生抑制のしくみ】
本研究では、R. aquaticaの気孔発生が抑制される仕組みを詳しく知るために、水没後にどのような遺伝子が働いているかを調べました。分析にあたっては、「RNA-seq解析※3」を利用しました(図3)。気孔は植物の重要な器官で、発生のメカニズムの詳細はすでに知られています。
これまでの研究で、若い葉の未分化な表皮細胞において、SPCHやMUTEなどの遺伝子が発現し、これによって気孔が分化することが分かっていました。我々の行った遺伝子発現解析の結果、R. aquaticaは水没1時間でこれらの遺伝子の発現を抑制し始め、24時間後にはほとんど発現しなくなることがわかりました。つまり、このことが水中での気孔の発生を抑制する直接の原因であったわけです。遺伝子発現のレベルでみると、水中葉への運命転換が1時間以内というのは、非常に早い応答であると言えます。
【水中葉への運命転換のしくみ】
それでは、R. aquaticaはどのように水没を感知しているのでしょうか?
我々の研究で、これには植物ホルモンである「エチレン※4」が決定的な役割を果たしていることがわかりました(図4)。エチレンは、気体として働く植物ホルモンです。RNA-seq解析の結果を見てみると、水没直後にエチレンにより制御される遺伝子の発現が大きく変動していました。そこで、気中で育てているR. aquaticaにエチレンを作用させたところ、気孔の発生が抑制されました。
気体のエチレンは、植物を水没させると逃げ場を失って植物の体内に蓄積されます。通常の陸上植物ではエチレンが作用しても気孔の発生は抑制されませんが、R. aquaticaは、進化の過程でエチレンのシグナル経路を再配線して(つなぎかえて)、水没時にSPCHやMUTEの遺伝子発現を抑制できるようになっているのです。また、今回の実験で我々は、R. aquaticaを暗所で水没させても気孔発生が抑制されないことを発見しました。R. aquaticaを、さまざまな波長の光が当たる環境においたところ、赤色光が当たっていない環境では水中葉は作られませんでした(図4)。また、赤色光がエチレンの合成を促進していることも、RNA-seq解析から裏付けられました。
以上から、我々は「赤色光の働きで合成されているエチレンは、植物が水没すると体内に蓄積し、SPCHとMUTEという気孔発生に必要な遺伝子の発現を抑制することで、水中葉への運命転換が素早く起こる」ことを突き止めました。
このように水中葉形成のメカニズムを分子レベルで明らかにしたのは、世界で初めての成果です(図5)。R. aquaticaはキャベツや菜種と同じアブラナ科植物です。比較的近縁の種であるモデル植物シロイヌナズナでは、エチレンや赤色光の応答経路の詳細が分子レベルで分かっています。しかしながら陸上植物であるシロイヌナズナは、水没してもエチレンや赤色光によってR. aquaticaのように気孔の形成が抑制されることはありません。したがって、シロイヌナズナの応答経路と今回の研究結果を比較することで、水陸両生植物の進化において、ホルモンや光の応答経路の再配線が重要な役割を果たしていたであろうと考えられます。
【今後の展開】
本研究で、水陸両生植物が水没などの環境変動にどのように応答しているのか、その一端を明らかにすることができました。近年、地球温暖化など大規模な環境変動が人類を脅かす危機として顕在化しています。今後の研究により、生物の、水位の変動が激しい環境への適応機構や、水環境への適応機構が明らかになれば、その成果は、陸上生態系の保護や、変動環境での農業生産性の向上などに寄与できるでしょう。
【責任著者コメント】
地面に根を張っている固着性の生活をしている植物は、環境の悪化などから逃げることができません。植物には静的なイメージがあるので、周囲の環境の変化に動的に応答しているようには感じないかもしれませんが、実は、移動できないからこそ、環境の変化に機敏に応答することで生き抜いているのです。本研究では、植物がどのように水没を感じ取り、気孔発生を抑制しているかを明らかにすることができました。今回の成果を取っ掛かりにして、植物がいかに巧みに環境変化に応答しているのか、さらに研究を進め、将来的に地球温暖化などへの対策にもつながる成果を出していきたいと思っています。
本研究は、文部科学省および日本学術振興会の科学研究費補助金(17H06476, 18H04844, 18H04787, 21H02513)、文部科学省の私立大学戦略的研究基盤形成支援事業(S1511023)の支援を受けて実施しました。
【用語説明】
※1 水陸両生植物
気中でも水中でも生育できる植物。気中と水中で葉の形や性質を大きく切り替える能力があるため、どちらの環境でも生育できる。
※2 Rorippa aquatica
アブラナ科イヌガラシ属の水陸両生植物。水没すると葉の形や性質を大きく変化させる。本研究室によりこの植物を使った研究が展開され、全ゲノム配列も解読されている。今回RNA-seq解析が実施できたのもゲノム配列情報を解読されていたからである。
※3 RNA-seq解析
細胞内のmRNAを取り出し、次世代シークエンサー※5を用いて網羅的に配列を解析することで、転写されている遺伝子(発現遺伝子)を同定する方法。解析対象の生物が持つ全遺伝子の発現情報を一度に得ることができる。
※4 エチレン
植物ホルモンの一種で、気体のホルモンである。りんごから出るエチレンがバナナの追熟に働くことで有名。
※5 次世代シークエンサー
DNAの配列を高速かつ大量に読むことができる配列解析装置。2000年代に次世代シークエンサーが登場したことで、それまでは非常に難しかった全ゲノム解析が研究室のレベルでできるようになり、生命科学に大きな革命をもたらした。
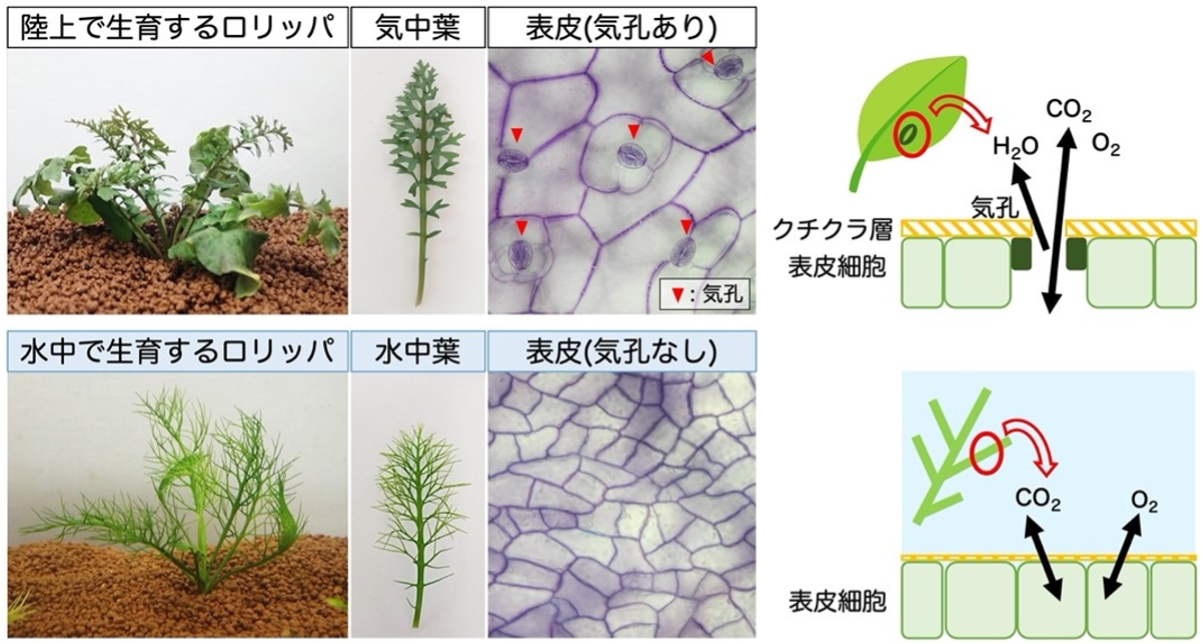
(上段)陸上で生育し、気中に葉(気中葉)を展開するR. aquatica。気中葉の表皮には多数の気孔が形成され、この孔を介して外部の空気とガス交換を行う。表皮細胞は厚いクチクラ層(ワックス)で乾燥から守られている。
(下段) 水中で生育し、水中に葉(水中葉)を展開するR. aquatica。水中葉の表皮には気孔が存在しない。表皮細胞が外部の水と直接ガス交換を行なっている。

(上段) 気中で生育した R. aquaticaを水没させる実験。青矢印は0日目の未成熟な葉を示し、7日目に成長している様子がわかる。水没しても新しい葉が形成されており、気中条件と変わらず成長を継続できている。
(下段) 未成熟な葉(上段:青矢印)の表皮を経時的に観察した。気中では葉の成長とともに多数の気孔が形成されるが、水中では気孔の形成が抑制されて新たに形成されないため、葉の成長とともに気孔の密度が低下する。
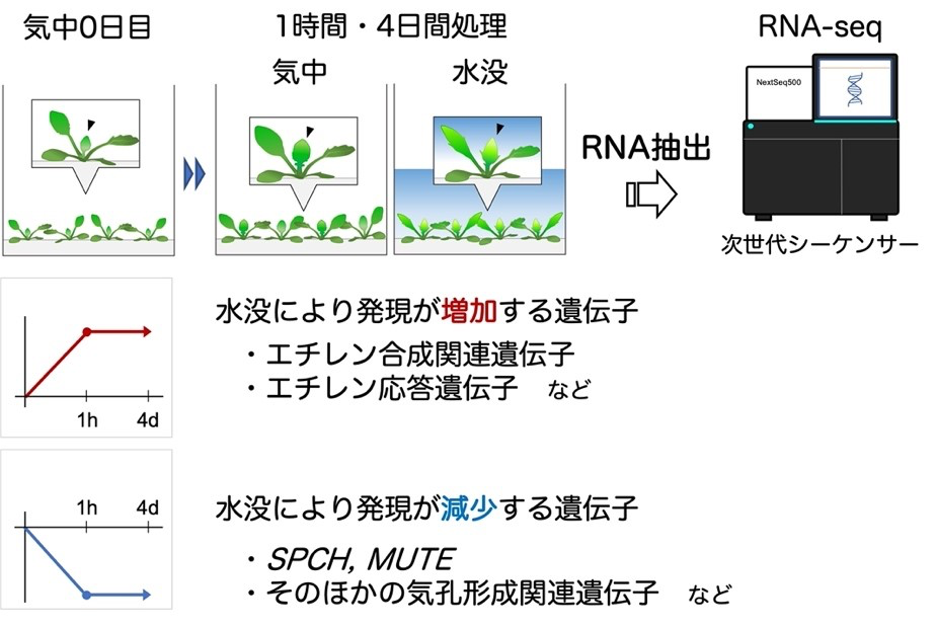
遺伝子はRNAとして転写されて機能する。気中で生育した植物を水没させ、未成熟な葉(図2:青矢印参照)からRNAを取り出して次世代シーケンサーで解析した。RNAの情報から遺伝子を特定し、R. aquaticaの全遺伝子の発現の動態を明らかにした。水没による遺伝子発現の変化に着目すると、エチレンに関連する遺伝子群が軒並み増加していた一方、気孔の形成に関わる遺伝子群は減少していた。特に、気孔形成に重要なSPCH, MUTEの2遺伝子は水没1時間で顕著に減少していることが示された。
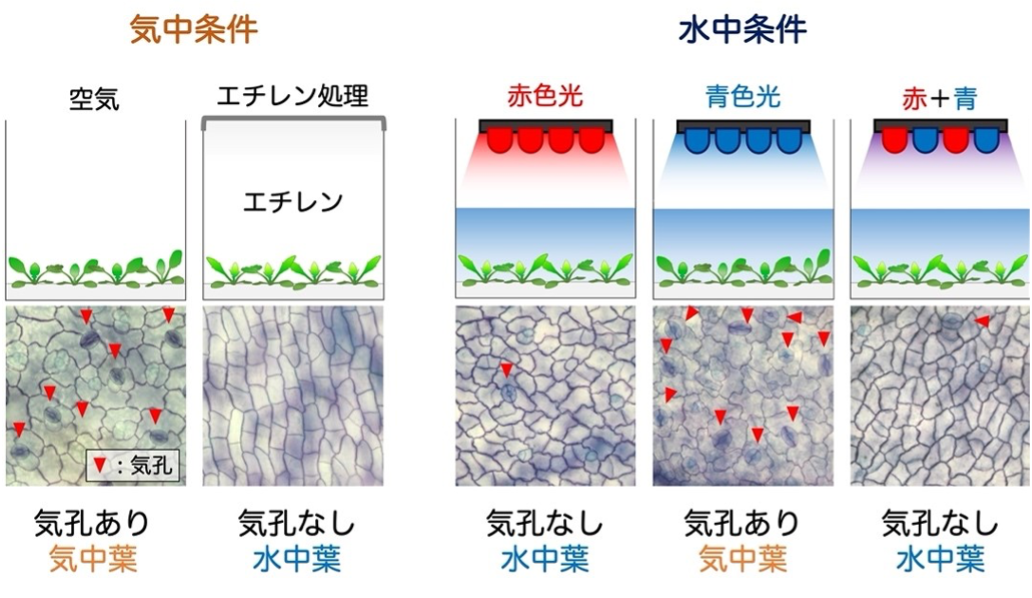
(左) エチレン処理実験。密閉したガラス容器内で空気にエチレンガスを添加して生育させると、気中であるにもかかわらず気孔のない水中葉が形成される。水没しなくてもエチレン単体で水没の効果を模倣でき、水中葉への切り替えにエチレンが重要な機能を果たしていることを示すことができた。
(右) 水没応答における光の効果。水没していても、暗所や青色光の照射下では気孔の形成を抑制できず気中葉が形成されてしまう。青に加えて赤色光も照射すると気孔形成が抑制されることから、水中葉の形成には赤色光が必要であることが明らかになった。自然環境の太陽光(実験室では白色光)には赤色成分も含まれているので水没に応答することができる。
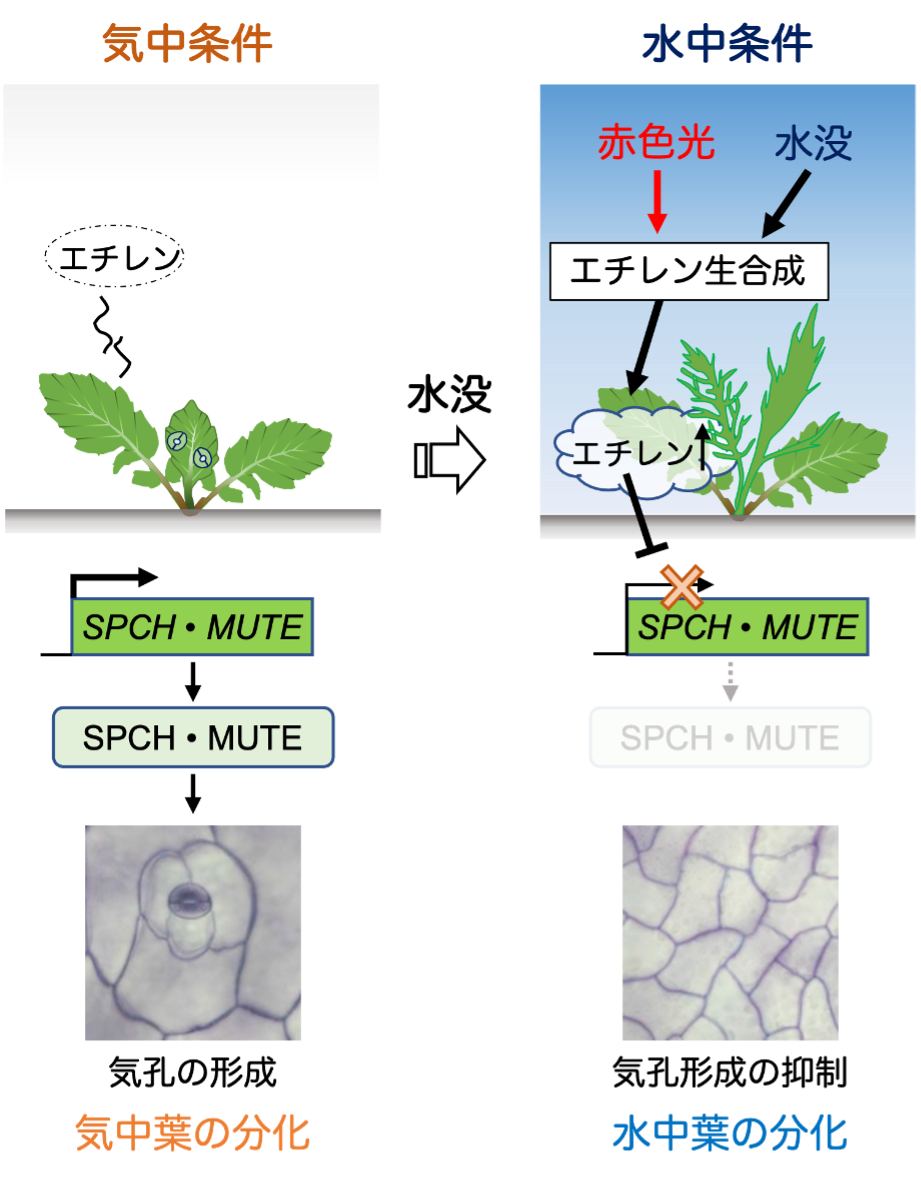
R. aquaticaはエチレンシグナルによりSPCH, MUTE遺伝子の発現を抑制できる新たな経路を獲得している。このような経路は近縁のモデル植物であるシロイヌナズナには存在しない。R. aquaticaは、陸上で生育する場合エチレンが体外の空気へと漏出するため一般的な陸上植物と同様に気孔を多数持つ気中葉を展開する。一方で水没すると、水中では体内にエチレンが蓄積することでエチレンシグナルが活性化される。 SPCH, MUTE遺伝子の発現が抑制されるため、気孔を持たない水中葉が分化する。さらに、水没を引き金にエチレンの合成が増加すること、これを赤色光が促進することで迅速な水没応答が実現している。
Information
| 論文タイトル | Rewiring of hormone and light response pathways underlies the inhibition of stomatal development in an amphibious plant Rorippa aquatica underwater (水陸両生植物Rorippa aquaticaの水没に応答した気孔分化抑制のメカニズムは、陸生祖先種の植物ホルモン応答経路と光応答経路を再配線(つなぎかえ)することで獲得された) |
|---|---|
| 著者 | Shuka Ikematsu, Tatsushi Umase, Mako Shiozaki, Sodai Nakayama, Fuko Noguchi, Tomoaki Sakamoto, Hongwei Hou, Gholamreza Gohari, Seisuke Kimura, Keiko U. Torii(池松 朱夏、馬瀬 樹志、塩崎 真子、中山 壮大、野口 楓子、Hongwei Hou、Gholamreza Gohari、木村 成介、鳥居 啓子) |
| 雑誌名 | Current Biology |
| DOI | 10.1016/j.cub.2022.12.064. |
| 発行年月 | 2023年1月 |